Gene Drift - Evolution Factor: Příčina, hodnota, příklady
Spolu s přirozenou selekcí existuje další faktor, který může ovlivnit nárůst obsahu mutantního genu. V některých případech může dokonce vyloučit normální alelu morph. Tento jev se nazývá genový drift v populaci. Podívejme se podrobněji na to, jaký je tento proces a jaké jsou jeho důsledky. 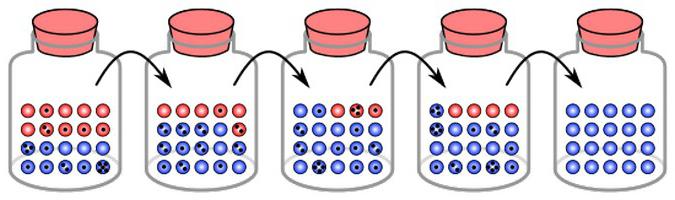
Obecné informace
Genetický drift, jehož příklady budou uvedeny v následujícím článku, jsou určité změny, které jsou zaznamenávány z generace na generaci. Předpokládá se, že tento jev má své vlastní mechanismy. Někteří výzkumní pracovníci se obávají, že v genofondu mnoha, ne-li všech, národů, množství nezvyklých genů, které se objevují, v současné době roste poměrně rychle. Určují dědičnou patologii, tvoří předpoklady pro rozvoj mnoha dalších onemocnění. Předpokládá se také, že patomorfóza (změna znaků) různých onemocnění, včetně onemocnění duševní povahy, je právě genovým driftem. Dotčený jev probíhá rychlým tempem. Výsledkem je číslo duševní poruchy vezme neznámé formy, stávají se nerozpoznatelnými ve srovnání s jejich popisem v klasických vydáních. Současně jsou významné změny zaznamenány přímo ve struktuře samotné psychiatrické morbidity. Například génový drift vymaže některé formy schizofrenie, které se objevily dříve. Namísto toho existují patologické stavy, které lze sotva určit moderními klasifikátory. 
Wrightova teorie
Náhodný genový drift byl studován pomocí matematických modelů. Použitím této zásady odvodil Wright teorii. On věřil, že klíčový význam driftu genu za konstantních podmínek je zaznamenán v malých skupinách. Stávají se homozygotními a variabilita se snižuje. Wright také věřil, že v důsledku změn ve skupinách se mohly vytvořit negativní dědičná charakteristika. V důsledku toho může celá populace zemřít bez přispění vývoje tohoto druhu. Současně výběr hraje velkou roli v mnoha skupinách. V tomto ohledu bude genetická variace v populaci opět nevýznamná. Postupně se skupina bude dobře přizpůsobovat podmínkám prostředí. Následující evoluční změny však budou záviset na výskytu příznivých mutací. Tyto procesy jsou poměrně pomalé. V tomto ohledu není vývoj velkých populací charakterizován vysokou rychlostí. U skupin mezilehlých hodnot je zaznamenána zvýšená variabilita. Současně dochází k náhodnému vzniku nových prospěšných genů, což zrychluje vývoj. 
Wrightovy závěry
Když se z populace ztratí jedna alela, může se objevit kvůli určité mutaci. Pokud je však druh rozdělen do několika skupin, v jednom z nich chybí jeden prvek, druhý v jiném, potom může gen přemísťovat od místa, kde to není, kde to není. Tak zůstane variabilita. Vzhledem k tomu Wright dospěl k závěru, že vývoj se objeví rychleji u těch druhů, které jsou rozděleny do četných nejrůznějších populací. Současně je mezi nimi možná migrace. Wright souhlasil přirozený výběr hraje velmi významnou roli. Nicméně, spolu s tímto výsledkem evoluce je drift genů. Identifikuje trvalé změny v pohledu. Navíc Wright věřil, že mnoho zvláštních rysů, které vznikly skrz drift, byly lhostejné a v některých případech dokonce škodlivé pro životaschopnost organismů. 
Výzkumní spory
K Wrightově teorii bylo několik názorů. Například Dobžánský věřil, že je bezvýznamné vzbudit otázku, který z faktorů je důležitější - přirozený výběr nebo genetický drift. Vysvětlil to svou interakcí. V podstatě jsou pravděpodobné následující situace:
- Pokud ve vývoji jednoho nebo druhého druhu, výběr zaujme vedoucí pozici, bude označena směrová změna genových frekvencí nebo stabilní stav. Ty budou určeny podmínkami prostředí.
- Pokud je v průběhu delší doby genový drift významnější, pak směrované změny nebudou způsobeny přírodním prostředím. Současně mohou nepříznivé znaky, i když se vyskytují v malých množstvích, šířit do skupiny dostatečně široce.
Mělo by však být poznamenáno, že samotný proces změny, stejně jako příčina genového driftu, není dnes dostatečně studován. V tomto ohledu neexistuje jediný a konkrétní názor na tento fenomén ve vědě.
Génový drift je faktor evoluce
Z důvodu změn je zaznamenána změna frekvence alel. K tomu dojde, dokud nedosáhnou stavu rovnováhy. To znamená, že génový drift je izolace jednoho prvku a fixace druhého. V různých skupinách se takové změny vyskytují nezávisle. V tomto ohledu jsou výsledky genetického driftu v různých populacích odlišné. Nakonec je některá skupina prvků pevná v některých, druhá v jiných. Génový drift proto na jedné straně vede ke snížení rozmanitosti. Současně však také způsobuje rozdíly mezi skupinami, odchylky z některých důvodů. To zase může sloužit jako základ pro speciaci. 
Poměr vlivu
V procesu vývoje genetický drift interaguje s dalšími faktory. Především je vztah založen na přirozeném výběru. Poměr příspěvků z těchto faktorů závisí na řadě okolností. Nejprve je určena intenzitou výběru. Druhou okolností je velikost skupiny. Takže pokud je intenzita a síla vysoká, náhodné procesy mají zanedbatelný vliv na dynamiku genetických kmitočtů. Současně, v malých skupinách s nevýznamnými rozdíly ve způsobilosti, je vliv změn neporovnatelně větší. V takových případech může být fixována méně adaptivní alela, zatímco adaptivnější bude ztracena.
Účinky změny
Jedním z hlavních výsledků genetického driftu je ochuzování rozmanitosti v rámci skupiny. To je způsobeno ztrátou některých alel a fixací ostatních. Proces mutace, naopak, naopak přispívá k obohacení genetické diverzity uvnitř populace. Kvůli mutaci se ztracená alela může objevit znovu a znovu. Vzhledem k tomu, že genetický drift je směrový proces, současně s poklesem intrapopulační diverzity se rozdíly mezi místními skupinami zvyšují. Proti tomuto fenoménu je migrace. Pokud tedy v jedné populaci je alela "A" fixní, a v druhé - "a", pak v rámci těchto skupin se znovu objeví rozmanitost. 
Konečný výsledek
Výsledkem genetického driftu bude úplné vyloučení jedné alely a konsolidace druhé. Čím častěji se ve skupině vyskytuje prvek, tím vyšší pravděpodobnost jeho fixace. Jak vyplývá z některých výpočtů, možnost vazby se rovná frekvenci alely v populaci.
Mutace
Vyskytují se v průměru s frekvencí 10-5 na gen na gametu na generaci. Všechny alely, které se nacházejí ve skupinách, se objevily v důsledku mutací. Čím je populace menší, tím nižší je pravděpodobnost, že každá generace bude mít alespoň jednoho jedince - nositele nové mutace. S počtem sto tisíc, každá skupina potomků s pravděpodobností blízkou jednotě má mutantní alelu. Nicméně jeho četnost v populaci, stejně jako možnost jeho fixace bude poměrně nízká. Pravděpodobnost, že stejná mutace se objeví ve stejné generaci, alespoň u jednoho jedince v populaci 10, je zanedbatelná. Avšak pokud se vyskytne u této populace, frekvence mutantní alely (1 z 20 alel), stejně jako šance na její fixaci, bude relativně vysoká. U velkých populací se objevuje nový prvek poměrně rychle. Současně je jeho fixace pomalá. Malé populace naopak očekávají dlouhou mutaci. Ale po jeho vzniku dochází k rychlejšímu upevnění. Z toho můžeme vyvodit následující závěr: možnost stanovení neutrálních alel závisí pouze na frekvenci mutačního výskytu. Současně velikost populace tento proces neovlivňuje.
Molekulární hodiny
Vzhledem k tomu, že frekvence výskytu neutrálních mutací u různých druhů je přibližně stejná, rychlost fixace by měla být přibližně stejná. Z toho vyplývá, že počet změn nahromaděných v jednom genu by měl být korelován s dobou nezávislého vývoje těchto druhů. Jinými slovy, čím déle bude čas od okamžiku oddělení dvou druhů od jednoho rodového původu, tím více rozlišují mutační substituce. Tento princip je základem metody molekulárně evolučních hodin. Toto definuje čas, který uplynul od okamžiku, kdy se předchozí generace různých systematických skupin začaly rozvíjet nezávisle, nezávisle na sobě.
Výzkum a studium Cukurkend
Tito dva američtí vědci zjistili, že počet rozdílů v aminokyselinové sekvenci v cytochromu a hemoglobinu u některých savčích druhů je vyšší, čím dříve jejich vývojové cesty se lišily. Následně byl tento model potvrzen velkým množstvím experimentálních dat. Materiál zahrnoval desítky různých genů a několik set druhů zvířat, mikroorganismů a rostlin. Ukázalo se, že průběh molekulárních hodin se provádí konstantní rychlostí. Tento objev je ve skutečnosti potvrzen danou teorií. Hodiny jsou kalibrovány odděleně pro každý gen. To je způsobeno skutečností, že frekvence výskytu neutrálních mutací v nich je odlišná. Za tímto účelem se vyhodnotí počet substitucí nahromaděných v daném genu v taxonech. Doba divergence je spolehlivě stanovena pomocí paleontologických dat. Po kalibraci molekulárních hodin je možné je dále používat. Zejména je s jejich pomocí snadné měřit čas, během kterého došlo k divergenci (divergence) mezi různými taxony. To je možné, i když jejich společný předchůdce nebyl dosud identifikován ve fosilních záznamů.