Dvojité oplodnění v rostlinách
Dvojité oplodnění v rostlinách má velký biologický význam. To bylo objeveno Navashin v roce 1898. Dále se blíže podíváme na to, jak dvojité hnojení se vyskytuje u rostlin. 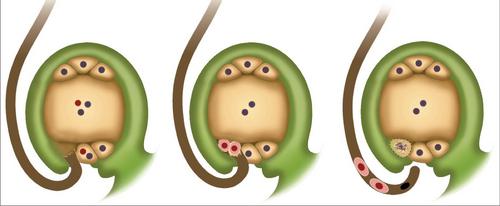
Biologický význam
Proces dvojího hnojení podporuje aktivní vývoj živinové tkáně. V tomto ohledu vajec neukládá materiál do budoucna. Toto vysvětluje svůj rychlý vývoj.
Dvojité oplodnění
Stručně řečeno, jev může být popsán následovně. Dvojité oplodnění v angiospermech spočívá v proniknutí dvou spermií do vaječníku. Jeden se slučuje s vejcem. To přispívá k začátku vývoje diploidního embrya. Druhá spermie se připojuje k centrální buňce. V důsledku toho se vytvoří triploidní prvek. Endosperm se objeví z této buňky. Je výživným materiálem pro vyvíjející se embryo.
Ad
Vývoj pylu
Dvojité oplodnění angiospermu začíná po vzniku vysoce redukované generace haploidů. To je reprezentováno gametophytes. Dvojité hnojení kvetinových rostlin podporuje klíčení pylů. Začíná napuštěním zrna a následnou tvorbou pylu. Prolomí sporoderm v nejtenčí sekci. Říká se tomu clona. Zvláštní látky jsou vypouštěny z hrotu pylu. Zmírňují tkáně sloupce a stigma. Kvůli tomu patří pylová trubice. Jak se vyvíjí a roste, do ní proniká sperma i jádro z vegetativní buňky. V drtivé většině případů pylová trubice proniká do nucellus (megasporangia) skrze mikropilot ovuly. To se dělá zřídka jiným způsobem. Po průniku do germinálního vaku se pylová trubice rozbije. Výsledkem je, že se celý jeho obsah nalévá uvnitř. Dvojité hnojení kvetinových rostlin pokračuje tvorbou diploidního zygota. To přispívá k prvnímu spermatu. Druhý prvek je spojen se sekundárním jádrem, které se nachází v centrální části embryového vaku. Vytvořené triploidní jádro se následně transformuje na endosperm. 
Tvorba buněk: obecné informace
Proces dvojího hnojení kvetinových rostlin se provádí pomocí speciálních zárodečných buněk. Jejich formování probíhá ve dvou fázích. První stupeň se nazývá sporogeneze, druhá je hematogeneze. V případě tvorby samčích buněk se tyto stavy označují jako mikrosporogeneze a mikrohematmatogeneze. Při vytváření prvků ženských genitálií se prefix změní na "mega" (nebo "makro"). Sporogeneze je založena na meióze. Toto je proces tvorby haploidních prvků. Meiosis, stejně jako mezi zástupci fauny, předchází množení buněk prostřednictvím mitotických dělení.
Ad
Produkce spermií
Primární formování samčích pohlavních elementů se provádí ve speciální tkáni. To se nazývá archesparial. V něm, jako důsledek mitóz, dochází k tvorbě četných elementů, mateřských pylových buněk. Pak vstoupí do meiózy. Kvůli dvěma meiotickým dělením se tvoří 4 haploidní mikrospory. Nějakou dobu leží v blízkosti a tvoří tetrad. Poté se rozpadají na zrna pylů - jednotlivé mikrospory. Každý z tvarovaných prvků začíná být pokryt dvěma skořápkami: vnější (exine) a vnitřní (intin). Pak začíná další fáze - mikrogametogeneze. To zase sestává ze dvou mitotických po sobě jdoucích divizí. Po vytvoření prvních dvou buněk: generativní a vegetativní. Následně první předá další divizi. V důsledku toho se tvoří dva samčí buňky - spermie. 
Makroporogeneze a megasporogeneze
V tkáních ovul se začíná oddělovat jeden nebo více obloukovitých elementů. Začínají rychle růst. Díky této aktivitě se stávají podstatně většími než ostatní buňky, které je obklopují v ovulech. Každý prvek arpspore je podroben rozdělení mitózou jeden, dva nebo vícekrát. V některých případech se buňka může okamžitě přeměnit na matku. Meióza se objevuje uvnitř. Výsledkem je vytvoření 4 haploidních buněk. Zpravidla se začíná rozvíjet největší z nich, čímž se stává germinální vak. Zbývající tři postupně degenerují. V této fázi se dokončí makrosporogeneze, začíná makrohematogeneze. Během ní se vyskytují mitotické dělení (většina angiospermů má tři). Cytokinéza nepředstavuje mitózy. V důsledku tří dělení se vytváří embryo vak s osmi jádry. Následně se izolují do nezávislých buněk. Tyto prvky jsou určitým způsobem distribuovány zárodečným vakem. Jedna z izolovaných buněk, která ve skutečnosti je vajíčková buňka, spolu s dalšími dvěma synergidy, se odehrává na mikropylonu, do něhož dochází k pronikání spermií. V tomto procesu hrají velmi významnou úlohu synergenty. Obsahují enzymy, které přispívají k rozpouštění membrán na pylu. Na opačné straně zárodečného vaku jsou umístěny další tři buňky. Jsou nazývány antipody. S pomocí těchto prvků se živiny přenášejí z vajíček na zárodečný vak. Zbývající dvě buňky se nacházejí v centrální části. Často se spojují. V důsledku jejich spojení se vytvoří diploidní centrální článek. Po dvojité oplodnění a spermie proniknou do vaječníku, jedna z nich, jak je uvedeno výše, se spojí s vajíčkovým buňkem. 
Vlastní pelenovou trubici
Dvojité oplodnění je doprovázeno jeho interakcí se sporofytovými tkáněmi. Je to docela specifické. Tento proces se řídí aktivitou chemických sloučenin. Zjistí se, že pokud se pyl vypláchne destilovanou vodou ztratí schopnost klíčit. Pokud je výsledný roztok koncentrovaný a poté zpracován, pak se opět zkompletuje. Vývin pelenové trubice po klíčení kontroluje tkáň pistil. Například u bavlny trvá její růst na vejce asi 12-18 hodin. Již po 6 hodinách je ovšem možné určit, na jaký ovul se pylová trubice dostane. To je pochopitelné, protože v ní začíná zničení synergie. V současné době není stanoveno, jak může rostlina směrovat vývoj trubice správným směrem a jak se synergent dozví o aproximaci. 
"Zakažte" sebepoškození
To je často pozorováno u kvetoucích rostlin. Tento jev má své vlastní charakteristiky. "Zákaz" sebepoškozování se projevuje ve skutečnosti, že sporofyt "identifikuje" svůj vlastní mužský hematofyt a nedovoluje mu účastnit se oplodnění. Současně v některých případech pyl plodu neprořídí svůj vlastní pyl. Růst trubice se však zpravidla začíná, ale následně se zastaví. Výsledkem je, že pyl nedosáhl vejce a v důsledku toho nedochází k dvojímu hnojení. Darwinovi byl tento fenomén zaznamenán. Takže našel v prvotřídní jarní květiny dvě formy. Některé z nich byly dlouhé stonky s krátkými tyčinkami. Jiné jsou krátké sloupce. V nich byly dlouhé vláknité vlákna. Květiny s krátkými stonky se liší ve velkém pylu (dvakrát tolik jako u ostatních). Současně jsou buňky v papilech stigmatu malé. Tyto příznaky jsou řízeny skupinou těsně tkaných genů. 
Receptory
Dvojité hnojení je účinné, když je pyl přenášen z jedné formy na druhou. Pro rozpoznání vlastních prvků jsou zodpovědné speciální molekuly receptoru. Jsou to složité sloučeniny karbohydrátů s bílkovinami. Bylo zjištěno, že formy divoké zelí, které nevytvářejí tyto receptorové molekuly v tkáních stigmatu, se samy opelejí. U běžných rostlin se objevují směsi sacharidů a bílkovin v den před otevřením květů. Pokud otevřete bud a zpracujete ho vlastním pylenem dva dny před jeho kvete, pak dojde k dvojímu hnojení. Pokud se tak stane den před otevřením, nebude to.
Ad
Alele
Je pozoruhodné, že v některých případech je "samo-neslučitelnost" pylu v rostlinách stanovena řadou několika prvků jednoho genu. Tento jev je podobný nekompatibilitě transplantace tkání u zvířat. Takové alely jsou označeny písmenem S. Počet v populaci těchto prvků může dosáhnout desítek nebo dokonce stovek. Například jestliže je genotyp rostliny produkující vajíčko s1s2 a s2s3 produkující pyl, při klíčovém opelení bude klíčení zaznamenáno pouze v 50% prachových částic. Jsou to ty, které nesou alelu s3. Pokud je několik desítek prvků, pak většina pylu bude normálně klíčit během křížení oříšků, zatímco samoopylení je zcela zabráněno. 
Na závěr
Na rozdíl od gymnospermů, které jsou charakterizovány vývojem dostatečně silného haploidního endospermu, bez ohledu na oplodnění, v angiospermech se tvoří tkáň pouze v tomto jediném případě. Vzhledem k obrovskému počtu generací bylo dosaženo značných úspor energie. Zvýšení stupně ploidy endospermu zřejmě přispívá k rychlejšímu růstu tkání v porovnání s diploidními vrstvami sporofytů.